2,4,5-TCPと2,4,6-TCPを分解する微生物の作成に関する研究
1. はじめに
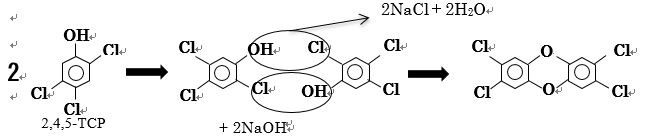
2,4,5-TCP (2,4,5-トリクロロフェノール) は殺菌剤や防カビ剤、2,4,6-TCPは殺菌剤や木材の防腐剤等に使われており、2,4,5-TCPはベトナム戦争で枯葉剤として使用された。クロロフェノール類は焼却灰・パルプ廃液等に存在し、アルカリ性でダイオキシンに変換される前駆体である。ダイオキシン等の環境ホルモンは、発がん性とともに極めて強い女性ホルモン性作用があり、乳がんや男性の精子数の減少への関与が指摘されており、人への健康に重大な影響を及ぼす。そこで、クロロフェノールが有害なダイオキシンになる前に、2,4,5-TCPや2,4,6-TCPの段階で分解させることを目的とした。
2,4,5-TCP (2,4,5-トリクロロフェノール) は殺菌剤や防カビ剤、2,4,6-TCPは殺菌剤や木材の防腐剤等に使われており、2,4,5-TCPはベトナム戦争で枯葉剤として使用された。クロロフェノール類は焼却灰・パルプ廃液等に存在し、アルカリ性でダイオキシンに変換される前駆体である。ダイオキシン等の環境ホルモンは、発がん性とともに極めて強い女性ホルモン性作用があり、乳がんや男性の精子数の減少への関与が指摘されており、人への健康に重大な影響を及ぼす。そこで、クロロフェノールが有害なダイオキシンになる前に、2,4,5-TCPや2,4,6-TCPの段階で分解させることを目的とした。
2. 実験方法
アルカリSDS法を用いて、分解遺伝子tftDを持つpTftDとpTTQ18(発現ベクター)のプラスミドを単離した後、EcoRIとHindIIIで制限酵素消化を行った。次に、切断したtftD遺伝子断片とpTTQ18を連結し、tftDのプロモーターをタンパク質の発現性の高いtacプロモーターへと変更した。その後、連結したプラスミドのタンパク質の発現をポリアクリルアミド電気泳動(SDS-PAGE)法を用いて確認した。その結果、tftDのタンパク質がわずかに発現していた。しかし、酵素を高発現させるためにプロモーターをT7に変更した。T7プロモーターを持つpET20bにtftDを連結した後、タンパク質の発現を確認したがタンパク質が発現していなかった。そこで、次にコールドショック発現ベクターpCold1に変更した。tftDをpCold1に連結し、E. coli BL21(DE3)を形質転換した後、タンパク質の発現を確認した。
アルカリSDS法を用いて、分解遺伝子tftDを持つpTftDとpTTQ18(発現ベクター)のプラスミドを単離した後、EcoRIとHindIIIで制限酵素消化を行った。次に、切断したtftD遺伝子断片とpTTQ18を連結し、tftDのプロモーターをタンパク質の発現性の高いtacプロモーターへと変更した。その後、連結したプラスミドのタンパク質の発現をポリアクリルアミド電気泳動(SDS-PAGE)法を用いて確認した。その結果、tftDのタンパク質がわずかに発現していた。しかし、酵素を高発現させるためにプロモーターをT7に変更した。T7プロモーターを持つpET20bにtftDを連結した後、タンパク質の発現を確認したがタンパク質が発現していなかった。そこで、次にコールドショック発現ベクターpCold1に変更した。tftDをpCold1に連結し、E. coli BL21(DE3)を形質転換した後、タンパク質の発現を確認した。
3. 実験結果及び考察
tftD遺伝子とpTTQ18を連結し、タンパク質の発現を確認した結果、目的のタンパク質の発現を確認できたが発現がhadAの発現に比較して1/10程度であった。そこで、タンパク質の高発現が予想されるT7プロモーターで発現させることを試みた。pET20bとtftD遺伝子を連結し、タンパク質の発現を確認したが、発現を確認できなかった。2つの制限酵素で切れるかを確認した結果、制限酵素の片方で切れていなかった為、pET20bのDNA配列が変化していると判断した。これは、pET20bが不安定な為、培養中に変化したと考えた。現在、pCold1にtftD遺伝子を連結し、tfdDタンパクの発現を調べている。
tftD遺伝子とpTTQ18を連結し、タンパク質の発現を確認した結果、目的のタンパク質の発現を確認できたが発現がhadAの発現に比較して1/10程度であった。そこで、タンパク質の高発現が予想されるT7プロモーターで発現させることを試みた。pET20bとtftD遺伝子を連結し、タンパク質の発現を確認したが、発現を確認できなかった。2つの制限酵素で切れるかを確認した結果、制限酵素の片方で切れていなかった為、pET20bのDNA配列が変化していると判断した。これは、pET20bが不安定な為、培養中に変化したと考えた。現在、pCold1にtftD遺伝子を連結し、tfdDタンパクの発現を調べている。